Takaväli 2. joulukuuta julkaistuista "arseenielämä" -lehdistä jatkuu edelleen. Osa kritiikistä on koskenut tiedettä, kun taas paljon enemmän kritiikkiä on ollut uutisten kattavuudesta ja myös siitä, kuinka NASA esitteli yleisöä tai ”kiusannut” yleisöä uutisilla käyttämällä sanoissa “astrobiologia” ja “maan ulkopuolinen elämä” ilmoitus tulevasta lehdistötilaisuudesta. Tänään, American Geophysical Union -konferenssissa, yksi ryhmän tutkijoista, Ron Oremland keskusteli laskeutumisesta uutisissa, ja annan siitä yleiskuvan siitä pian. Noin samaan aikaan tiederyhmä julkaisi lausunnon ja joitain usein kysyttyjä kysymyksiä tiedekirjasta. Alla on tämä lausunto ja tiedetiimin toimittamat tiedot.
Vastaus kysymyksiin, jotka liittyvät tieteelliseen artikkeliin "Bakteeri, joka voi kasvaa käyttämällä arseenia fosforin sijasta"
- Kuten 16. joulukuuta 2010-
Science-lehden 2. joulukuuta 2010 julkaisema tutkimusartikkeli tarjosi useita todisteita, jotka viittaavat yhdessä siihen, että Kalifornian Monojärvestä eristetty bakteeri voi korvata arseenin pienellä prosentilla fosforia ja ylläpitää sen kasvua.
Tämä löytö oli yllättävää, koska kuusi elementtiä - hiili, happi, vety, typpi, rikki ja fosfori - muodostavat suurimman osan elävän aineen orgaanisista molekyyleistä, mukaan lukien nukleiinihapot, proteiinit ja lipidit. Siksi tutkijat, jotka eivät ole sidoksissa tutkimusryhmään, ovat kysyneet tutkimuksesta asianmukaisesti haastavia kysymyksiä.
Tutkimusjulkaisun keskeinen tarkoitus on edistää tiedettä esittämällä mielenkiintoista tietoa ja ehdottamalla testattavia hypoteeseja. Ymmärrettävästi yllättävimmillä havainnoilla on taipumus saada aikaan tiedeyhteisöltä eniten vastauksia ja tarkastuksia. Alkuperäisen tutkimuksen julkaisemisen jälkeiset vastaukset ja ponnistelut tulosten testaamiseksi ja toistamiseksi, etenkin odottamattomien havaintojen tapauksessa, ovat olennainen mekanismi tieteellisen tiedon edistämiselle.
Tiedetoimittajat ovat nyt vastaanottaneet useita teknisiä kommentteja ja kirjeitä, joissa vastataan Felisa Wolfe-Simonin ja hänen kollegoidensa artikkeliin ”Bakteri, joka voi kasvaa käyttämällä arseenia fosforin sijasta”. Kommentit ja vastaukset tarkistetaan, ja julkaisemme ne tulevassa Science-lehdessä.
Samaan aikaan pyrkiessään edistämään julkista ymmärrystä teoksesta, tutkimusartikkeli ja siihen liittyvä uutinen on asetettu vapaasti yleisön saataville seuraavan kuukauden ajan Science-verkkosivuston kautta. Nämä artikkelit löytyvät verkosta täältä:
Wolfe-Simon-joukkue teoreettisesti, että jotkut bakteerit saattavat pystyä käyttämään arseenia tai sietämään jonkin verran fosforin korvaamista orgaanisissa molekyyleissä, keräsivät mikrobit arseenirikkaasta Monojärvestä ja vieroittivat sitten ne vähitellen fosforista syöttämällä heille sijaan arseenia. Ryhmä on ilmoittanut ryhtyneensä toimiin fosforin saastumisen estämiseksi. He päättelivät, että heidän todisteidensa mukaan arseeni oli korvannut pienen osan DNA: n fosforista.
Kirjailijat ovat kuvanneet erityyppisiä todisteita, mukaan lukien:
* Induktiivisesti kytketty plasmamassaspektrometria.
Kirjoittajat kertoivat, että nämä tulokset paljastivat, että arseenia oli bakteerisoluissa, mikä viittasi siihen, että se ei olisi pelkästään solujen ulkopuolelle tarttunut epäpuhtaus;
* Arseenin radioaktiiviset merkinnät.
Wolfe-Simonin joukkue sanoi, että näiden todisteiden ansiosta he havaitsivat normaalisti myrkyllisen aineen solujen proteiini-, lipidi-, nukleiinihappo- ja metaboliittifraktioissa, mikä viittaa siihen, että se on otettu molekyyleihin, jotka muodostavat kunkin fraktion.
* DNA: n korkearesoluutioinen sekundaarinen ionimassaspektrometria sen jälkeen, kun se oli erotettu bakteereista.
Kirjoittajat kertoivat, että nämä todisteet viittasivat siihen, että eristetty DNA sisälsi edelleen arseenia.
* Korkean intensiteetin (synkrotroninen) röntgenanalyysi.
Tämän todisteen perusteella kirjoittajat päättelivät, että bakteerien arseeni näytti korvaavan fosfaatit DNA: ssa ja muissa molekyyleissä.
Havainnoista havainnoista on yleensä keskitytty siihen, olivatko bakteerit todella sisällyttäneet arseenia DNA: hon ja ovatko mikrobit lopettaneet fosforin käytön kokonaan. Vaikka ryhmä pyrkii vastaamaan kysymyksiin vertaisarvioidun prosessin kautta, Felisa Wolfe-Simon ja Ron Oremland ovat toimittaneet täällä julkisen palveluna joitain lisätietoja ja selventäneet tietojaan ja menettelyjään. Tiede korostaa, että näitä vastauksia ei ole vertaisarvioitu; ne toimitetaan tekijöiden puolesta vain julkisena tietopalveluna, kun taas tiedelle lähetettyihin kommentteihin annettujen vastausten muodollisempi tarkastelu jatkuu.
Alustavat kysymykset ja vastaukset
Kysymys: Jotkut ihmiset ovat kysyneet, puhdistettiinko DNA: n tekniikka riittävästi geelielektroforeesilla, jotta se voitaisiin erottaa muista molekyyleistä. Katsotko, että tämä on pätevä huolenaihe?
Vastaus:
DNA: n uutto- ja puhdistusprotokollamme alkaa pestyistä soluista, jotka on pelletoitu väliaineista. Näille suoritetaan sitten tavanomainen DNA-uuttoprotokolla, joka sisälsi useita fenolikloroformivaiheita epäpuhtauksien poistamiseksi, mukaan lukien kaikki sisällyttämättömät arsenaatit (As). Tämän jälkeen DNA elektroforeesoitiin edelleen erottaen DNA epäpuhtauksista. Mahdolliset väliaineen jäännökset olisi poistettu pesemällä solut ennen uuttamista ja jakamalla vesifaasiin uuton 3 fenoli: kloroformi-vaiheen aikana. Jos As sisällytettäisiin lipidiin tai proteiiniin, se olisi jakautunut fenoli-, fenoli: kloroformi- tai kloroformifraktioihin. Lisäksi tällä tavalla muista näytteistä uutettua DNA: ta käytettiin menestyksekkäästi myös lisäanalyyseissä, mukaan lukien PCR, jotka vaativat erittäin puhdistettua DNA: ta.
NanoSIMS: n mittaama arseeni geelinauhalla on yhdenmukainen muiden mittausten ja toisen todistusaineiston kanssa.
Radioaktiivisella leimatulla 73AsO43- kokeilumme osoitti, että solupelettiin liittyvästä kokonaismäärästä 11,0% ± 0,1% liittyi DNA / RNA-fraktioon. Tämä osoitti, että meidän pitäisi odottaa olevan jonkin verran arsenaattia nukleiinihappoihin liittyvästä kokonaispoolista. Näiden tietojen tulkitsemiseksi yhdistämme tulkinnan EXAFS-todisteisiin, jotka viittaavat siihen, että solunsisäinen arseeni oli As (V) sitoutunut C: ään eikä se ollut vapaa liuoksessa ionina. Tämä viittaa sellaiseen orgaaniseen molekyyliin, jonka sidosetäisyydet ovat sopusoinnussa kemiallisen ympäristön kanssa, joka on analoginen fosfaatin kanssa (kuviot 3A, S3-taulukko “sidospituudet”). Tuemme edelleen tulkintamme edellisistä kahdesta analyysistä, käytimme NanoSIMS: n kolmatta todistussarjaa, täysin erilaista tekniikkaa kuin kaksi muuta. Löydämme geelikaistaan liittyvää alkuaineen arseenia (mitattuna NanoSIMS), joka on enemmän kuin kaksi kertaa tausta geelissä. Edellä esitetyn keskustelun perusteella emme mielestäni ole aiheellista huolta.
Kysymys: Toiset ovat väittäneet, että arsenaattiin sidotun DNA: n olisi pitänyt hajoa nopeasti, kun se altistetaan vedelle. Voisitko puhua tähän?
Vastaus:
Meillä ei ole tietoa sellaisista tutkimuksista, jotka koskisivat pitkien ketjujen polyestereihin tai arsenaatin nukleotididi- tai triestereihin sitoutuneita arsenaattia. Julkaistut tutkimukset ovat osoittaneet, että yksinkertaisilla arseeniestereillä on paljon korkeammat hydrolyysinopeudet kuin fosfaattiestereillä (1-3). Tähän päivään mennessä julkaistuissa kokeissa on tarkasteltu erityisesti arsenaatin alkyylitriiestereiden vaihtoa tai hydrolyysiä [Eqn. 1] ja arseniitin alkyylidi-esterit [Eqn. 2]:
OAs (OR) 3 + H20? OAs (OH) (OR) 2 + ROH [1]
OAs (OH) (OR) 2 + H20? OAs (OH) 2 (OR) + ROH [2]
jossa R = metyyli, etyyli, n-pentyyli ja isopropyyli. Viite 2 osoitti, että näiden yksinkertaisten arsenaatin alkyyliesterien hydrolyysinopeudet laskivat kasvaessa alkyylisubstituentin (metyyli> etyyli> n-pentyyli> isopropyyli) hiiliketjun pituutta (monimutkaisuutta). Arsenaattiin sidottujen nukleotidien tai muiden biologisesti merkityksellisten osien hydrolyysinopeuksille ei ole tehty työtä.
Jos hydrolyyttisen nopeuden trendi, joka on ilmoitettu viitteessä 2 2 jatkaa suurempien painoisten orgaanisten aineiden, kuten sellaisten, joita löytyy biomolekyyleistä, on ajateltavissa, että arsenaattiin sidotut biopolymeerit saattavat olla vastustuskykyisempiä hydrolyysille kuin aiemmin ajateltiin. Viitteissä Refs. Kuviot 1-3 ovat suhteellisen joustavia ja voivat helposti omaksua ihanteellisen geometrian veden hyökätäkseen arsenoesterisidoksen kanssa. Suurten biomolekyylien arsenaattiesterit ovat kuitenkin todennäköisesti steerisemmin estyneitä, mikä johtaa hitaampaan hydrolyysinopeuteen.
Tämän tyyppinen steerinen reaktionopeuden rajoitus vastaa monenlaisia nopeuksia, jotka nähdään joidenkin fosfaattiin sidottujen nukleotidien käyttäytymisessä. Pienissä ribotsyymeissä fosfodiesterisidokset katalyysipaikassa voidaan hydrolysoida kymmenien sekuntien luokkaa (kemiallisella nopeudella 1 s-1). Tämä nopeuden parannus saavutetaan orientoimalla sidos nukleofiilin (vierekkäisen 2'-hydroksyyliryhmän) in-line-hyökkäykseen. Lisäksi automaattisen hajoamisen kuviot ovat yhdenmukaisia tietyn emäskoostumuksen kanssa. Toisaalta fosfodiesterisidosten hydrolyysinopeudet RNA: n A-muodon duplekseissa ovat monen suuruusluokkaa hitaampia, koska nämä liitokset eivät pääse helposti hydrolyysin edellyttämään geometriaan.
DNA: n nopeudet voivat olla paljon hitaampia kuin malliyhdisteet, koska kierre asettaa selkärankaan geometriset rajoitukset.
Arsenaattiin sidottujen biopolymeerien hydrolyysin kinetiikka on selvästi alue, jolla on enemmän tutkimusta perusteltua.
Kysymys: Onko mahdollista, että kasvualustasi suolat olisivat voineet tarjota tarpeeksi fosforin jälkeä bakteerien ylläpitämiseksi?
Vastaus:
Taulukon S1 tiedot ja näytteiden merkinnät ovat aiheuttaneet hämmennystä. Jokaisen kokeen selventämiseksi tehtiin yksi erä keinotekoista Mono Lake -vettä seuraavalla formulaatiolla: AML60-suolat, ei P, ei As, ei glukoosi, ei vitamiinit. Taulukko S1 näyttää esimerkkejä alkuainefosforin (~ 3 uM) ja arsenaatin ICPMS-mittauksista, jotka on tehty tällä formulaatiolla ennen mahdollisia lisäyksiä. Sitten lisäsimme glukoosia ja vitamiineja kaikkiin kolmeen hoitoon ja joko As + + -hoitoihin tai P + + -hoitoihin. Elatusaineelle tehdyt P-mittaukset sakkaroosin ja vitamiinien lisäämisen ja As: n lisäyksen jälkeen olivat myös ~ 3 uM tässä erässä. Siksi oli selvää, että mikä tahansa mitattu P-epäpuhtaus (~ 3 uM, tämä oli suuri alue) tuli tärkeimpien suolojen kanssa ja että kaikki kokeet sisälsivät identtistä P-taustaa (mukaan lukien mikä tahansa viljelykierrossa mukana oleva P).
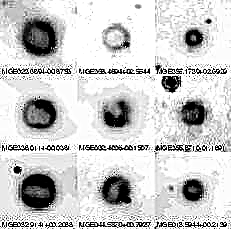
Science-lehdessä esitetään tulokset yhdestä monesta toistettujen kokeiden kokeesta, joka ei osoita solujen kasvua elatusaineissa ilman lisättyä arsenaattia tai fosfaattia (kuva 1). Nämä tiedot osoittavat selvästi, että kanta GFAJ-1 ei kyennyt käyttämään 3 uM P: tä tukemaan lisäkasvua ilman arsenaattia. Lisäksi + As / -P-kasvatetuille soluille määritetty solunsisäinen P-pitoisuus ei ollut riittävä tukemaan P: n täydellistä vaatimusta solun toiminnan kannalta.
Huomautus viljelystä: Kaikki kokeet aloitettiin inokulaateilla jatkuvista + As / -P-olosuhteista. Ennen kokeita soluja oli kasvatettu pitkäaikaisesti useita sukupolvia yhdestä pesäkkeestä, joka oli kasvatettu kiinteässä väliaineessa ilman lisättyä fosfaattia. Ennen tätä niitä kasvatettiin rikastuttajana yli 10 siirrokseen ja aina uuteen kasvualustaan, joka oli + As / -P. Siksi mielestämme P: n siirtämistä ei ole tapahtunut merkittävästi. Väitämme myös, että solun P-solua ei olisi ollut tarpeeksi tukemaan lisäkasvua, joka perustuu P: n sisäiseen kierrätysvarastoon.
Kysymys: Onko jotain muuta, jonka haluat yleisön ymmärtävän tutkimuksestasi tai tieteellisestä prosessista?
Vastaus: Meille kaikille, koko joukkueellemme, millaista tämä oli, oli mahdotonta kuvitella. Olemme ryhmä tutkijoita, jotka kokoontuivat ratkaisemaan todella mielenkiintoisen ongelman. Me kukin käytimme kykyjämme teknisestä osaamisesta henkiseen keskusteluun objektiivisen määrittämiseksi, mitä tarkalleen tapahtui kokeiluissamme. Tunnustimme vapaasti lehdissä ja lehdissä, että meidän ja koko joukon muiden tutkijoiden oli tehtävä paljon, paljon enemmän työtä. Lehdistötilaisuudessa oli mukana jopa tekninen asiantuntija tohtori Steven Benner, joka ilmaisi joitain huolenaiheisiin, joihin me jo vastasimme. Osa syytämme tuoda tämä työ yhteisöön oli luoda henkiset ja tekniset yhteydet yhteistyön lisäämiseksi vastaamaan moniin pysyviin kysymyksiin. Olimme läpinäkyviä tietojemme kanssa ja osoitimme jokaisen pisteen ja mielenkiintoisen tuloksen. Lehtymme päätelmät perustuvat siihen, mikä oli mielestämme kaikkein haastavin tapa tulkita koesarjaa, jossa yksikään koe ei pysty vastaamaan suureen kysymykseen. "Voisiko mikrobi käyttää arseenia fosforin sijasta ylläpitää sen kasvua?" Paras tiede avaa uusia kysymyksiä meille yhteisöksi ja herättää suuren yleisön mielenkiinnon ja mielikuvituksen. Kommunikoijina ja tieteen edustajana katsomme, että uusien ideoiden tukeminen tiedolla on kriittistä, mutta samalla myös uusien ideoiden tuottamiseksi muiden ajattelemiseksi ja kykyjensä hyödyntämiseksi.
Odotamme innolla yhteistyötä muiden tutkijoiden kanssa joko suoraan tai asettamalla solut vapaasti saataville ja toimittamalla DNA-näytteitä sopiville asiantuntijoille niiden analyysejä varten pyrkiessämme saamaan enemmän tietoa tästä kiehtovasta löytöstä.
Viitteet
1. T. G. Richmond, J. R. Johnson, J. O. Edwards, P. H. Rieger, Aust. J. Chem. 30, 1187 (1977).
2. C. D. Baer, J. Rieger, Inorg. 20, 905 (1981).
3. J.-M. Käsityöt, härkä. Soc. Chim. Fr. 14, 99 (1870).
4. Lagunas, D. Pestana, J. Diez-Masa, Biochemistry 23, 955 (1984).
Lähde: Felisa Wolf-Simonin verkkosivusto, Iron Lisa